Author: Kavya Rajagopalan
Mentors: Michael Brenner, Erik Winfree, Chrisy Xiyu Du, Ofer Kimchi, Agnese Curatolo
Editor: Wesley Huang
Introduction
Over the last few decades, nanotechnology, the construction of extremely small devices, has become a rapidly developing field. Nucleic acids can be used to successfully synthesize various 2-D and 3-D nanoscale structures, ranging from circuits to autonomous motors (Srinivas et al 2013). The applications of this research are vast: researchers are working to develop molecular machines, DNA computers, and various other nanoscale devices. This technique of utilizing DNA to engineer nanoscale objects is known as DNA Origami and serves as a valuable method for controlling, modifying, and storing essential computational information, which is critical in algorithmic self-assembly: a field of study on programming molecules to manipulate themselves.
Despite its many computational uses, DNA Origami is far from a perfect art. This process currently poses several limitations that prevent it from being a suitable candidate for constructing many nanoscale structures. For example, DNA strands take a long time to assemble, meaning only small nanoscale structures can be successfully constructed. Additionally, DNA’s alphabet is limited to A, T, G, and C, so the resulting models are relatively simple and do not possess the complexity necessary for constructing higher order devices (Srinivas et al 2013).
Given these limitations in the current DNA origami model, the Brenner group, in collaboration with the McEuen and Cohen group at Cornell, have proposed a novel solution, which suggests that specifically designed magnetic panels (known as a “magnetic handshake system”) can be substituted in place of traditional DNA base pairs. In theory, if this model could emulate all the characteristic behaviors of DNA, it would be the perfect solution to circumventing the same constraints that make the DNA model so ineffective. Not only would it yield a more controlled and faster assembly process, but with more options for distinct base pairs, it would also result in a more complex language (Niu et al 2019). Each disk has a unique magnetic configuration which corresponds to a “letter” in a traditional DNA strand (Figure 1A). The physical binding of these magnetic DNA strands will look and behave almost identically to a generic DNA strand (Figure 1B).
In order to successfully construct such a model, a variety of DNA behaviors must be well understood. One such characteristic behavior is oscillation, which describes the fluctuating nature of a specific DNA strand’s concentration when two or more distinct DNA molecules interact. Thus, being able to control oscillations is particularly useful as it allows us to adjust how multiple DNA molecules behave in each other’s presence and how we may want to construct a specific DNA-nanostructure.
Thus, being able to determine how oscillations can be translated into this magnetic handshake system is crucial to further developing this model. To ultimately achieve this goal, it is necessary to address the following key questions:
(1) How exactly do DNA strands create oscillations?
(2) How can we use the magnetic system to fully control the speed and behavior of an oscillation?

Figure 1. A. Four different magnetic panels and their corresponding binding panels. Each panel has a distinct magnetic dipole configuration, which in turn corresponds to a different binding energy that as the users we have the ability to encode for. B. An experiment in which two magnetic “DNA strands” begin to bind. Reprinted from “Magnetic handshake materials as a scale-invariant platform for programmed self-assembly,” by R.Niu, 2019, PNAS.
Objectives and Outcomes
The goals of this project are as follows:
- Identify how our magnetic system can replicate DNA oscillations
- Assess how users can gain complete control of the oscillation speed
When we construct this panel system, we only know two parameters for certain: the binding energies and the length of our strand. So, to achieve our goals, we will undertake the following steps:
- Identify some critical mechanism that both DNA and oscillations depend on, while characterizing it as a function of strand length and binding energies.
- Take some arbitrary oscillator and write it as a series of DNA reactions.
- Using step 1, identify how the rate of our oscillator is dependent on strand length and binding energy.
- Create a magnetic system, and use the binding energies and strand length to create an oscillation with the behavioral patterns and features that we are looking for.
It has previously been demonstrated that constructing both oscillations and a variety of other DNA nanoscale devices is highly dependent on a process known as Toehold Mediated Strand Displacement (TMSD). TMSD is a mechanism used between multiple DNA strands to control the reaction kinetics of systems that require DNA (Srinivas et al 2013). Essentially, this process serves as a remote, both activating and regulating the speed at which DNA-based nanoscale structures assemble. In TMSD, an invading strand binds to an unbound “toehold” region. It undergoes a random walk and displaces the incumbent strand in order to create a new duplex (Figures 2, 3).
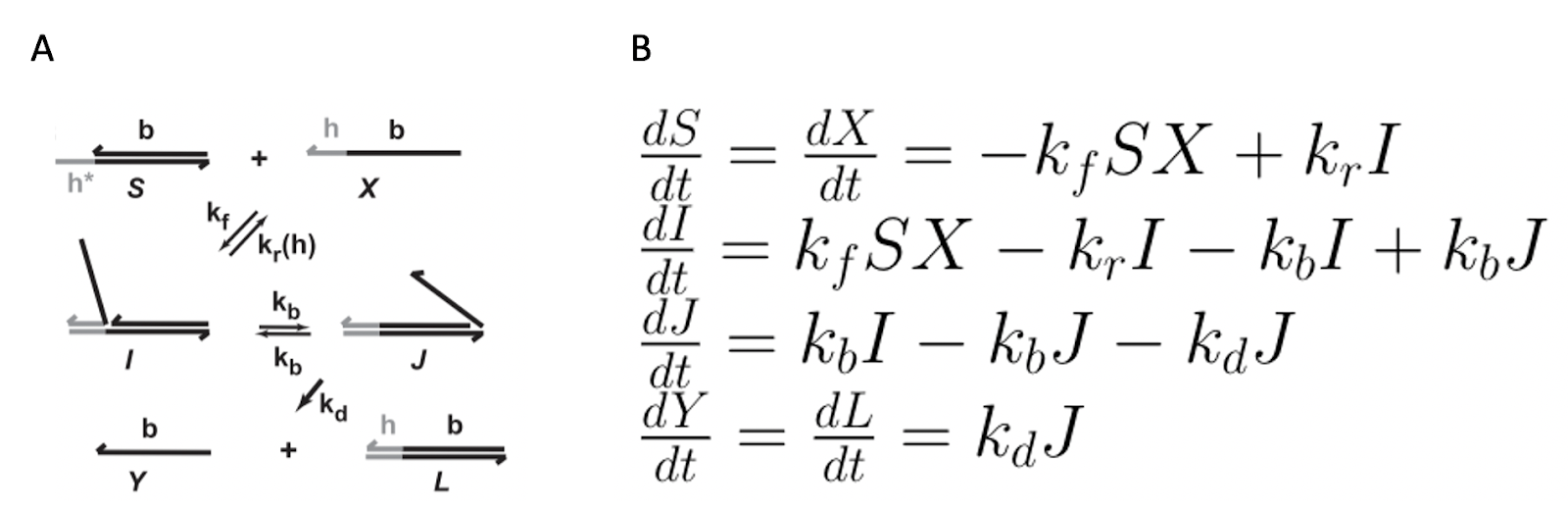
Figure 2. A. Mechanism of TMSD. The strand displacement step (I → J), appears as one single reaction; however, this step is a random walk, meaning it is composed of multiple random sub-steps. Reprinted with permission from “On the biophysics and kinetics of toehold-mediated DNA strand displacement,” by N. Srinivas, 2013. B. Change in concentration of species in TMSD as a reaction proceeds.
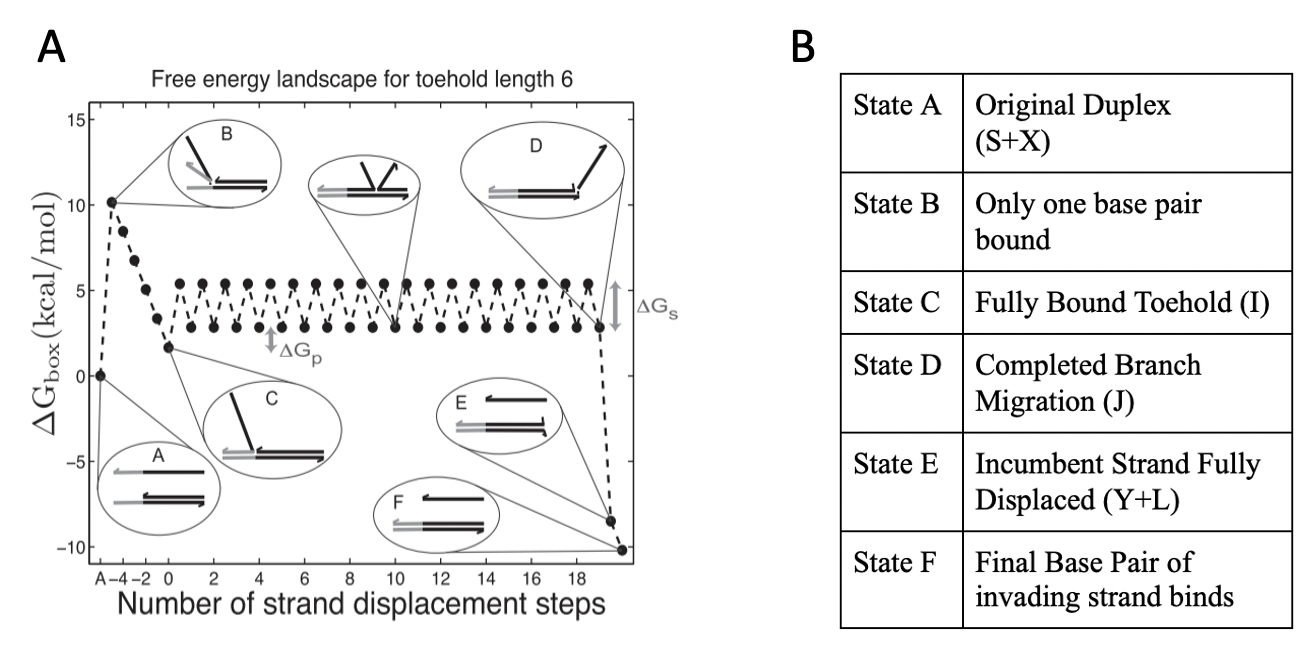
Figure 3. A. Free energy penalties associated with each step of TMSD. Each spike indicates a loss or gain in free energy as a function of strand displacement steps. Notice that steps C → D exhibits a sawtooth pattern, indicating that base pairs constantly fluctuate between binding and unbinding, meaning this step is a random walk. Reprinted with permission from “On the biophysics and kinetics of toehold-mediated DNA strand displacement, ” by N. Srinivas, 2013. B. States shown in A, and corresponding description of the process.
Approach and Methodology
To determine the rate of a singular TMSD reaction, it is necessary to identify the rates of each individual step from Figure 2 and Figure 3. TMSD first requires that an invading strand bind at the toehold of a template strand. The invading strand will displace the incumbent, or already-bound, strand and completely attach to the template. Identifying the rates of most of these steps is fairly straightforward; however, the strand displacement step (C ⇄ D in Fig 3, I ⇄ J in Fig 2) is frequently treated as a free parameter. In reality, this step is far more complex since it is a random walk that consists of a unimolecular step (the base pairs bind: the “up” steps in the sawtooth pattern) and a bimolecular step (the base pairs unbind: the “down” step in the sawtooth pattern). Each of these processes has a different rate that occurs the same number of times throughout an entire strand displacement reaction. In order to find the rate of this random walk, we generated a model for identifying the relationship between the number of unimolecular and bimolecular steps in the sawtooth as a function of the toehold length and binding energy. This model is intended for strand lengths ranging anywhere from 10 to 20 base pairs long, since these are the only viable lengths that can be used in the magnetic system as most DNA strands only undergo TMSD at this length. After thousands of simulations, we arrive at the following simple relationship:
Steps = 60.3* (strand displacement length) – 437
Since this process is a single order rate reaction, identifying the rate of this step (kb) became clear enough once we arrived at the above relationship. However, finding the rate of the overall TMSD reaction meant identifying how the overall effective rate of a TMSD reaction is dependent on its sub-rates. The final conclusion we arrive at is that a TMSD reaction is described by:
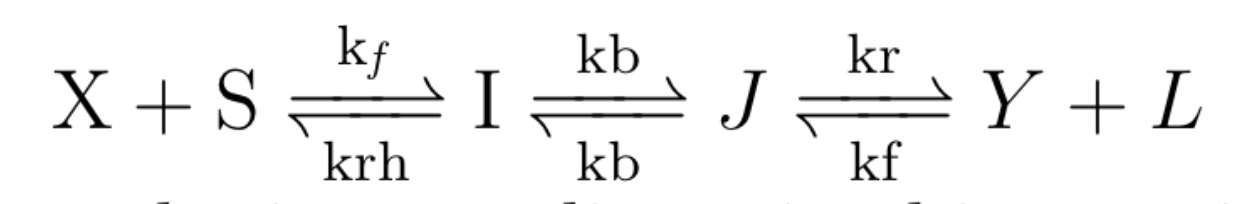
With some algebraic manipulation, we arrive at the following equation which is validated in Srinivas et al 2009:
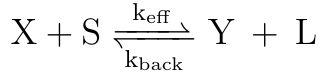
After determining what the values of kr, kf, kr(h) and kb(the forwards and backwards rates for each step in TMSD) would be, we plugged these individual rates into the equation for keff, and identified the relationship between the rate of TMSD and the toehold length at a given binding energy (Figure 4).
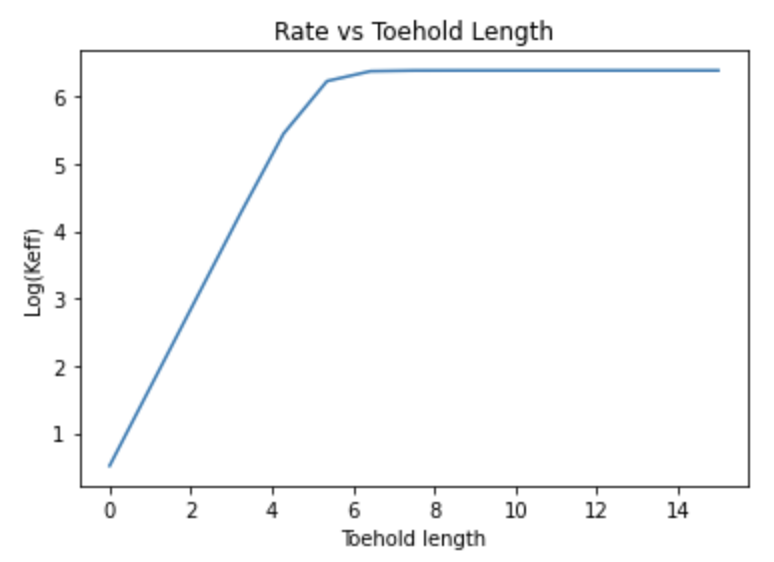
Figure 4. Relationship between rate of TMSD and toehold length. When we alter the individual rates for each step in TMSD, or the initial concentration of the DNA strands which initiate TMSD, we notice that the toehold length corresponding to when the curve begins to plateau also varies. However, the general shape of the curve remains the same.
This outcome indicates that an oscillation, which is composed of a series of DNA reactions, can also be written as a series of strand displacement reactions. Consequently, if we understand how the overall rate of an oscillation is dependent on the rates of the individual strand displacement reactions, we can identify the overall reaction rate of an oscillation given a specific binding energy and strand length.
Results
We focused on characterizing the effective rate of the Rock Paper Scissor (RPS) oscillator. The RPS Oscillator has been extensively studied, and many other oscillators can be constructed by slightly modifying the RPS oscillator. In an ideal scenario, the RPS Oscillator is dictated by the following reactions:
A + B → 2B at rate k1 (module 1)
B + C → 2C at rate k2 (module 2)
A + C→ 2A at rate k3 (module 3)
When we modify the values of k1, k2, k3, we are left with two plots where the periods of oscillation vary dramatically, suggesting that these individual rates are just as influential in the oscillations as we suspected (Figure 5).
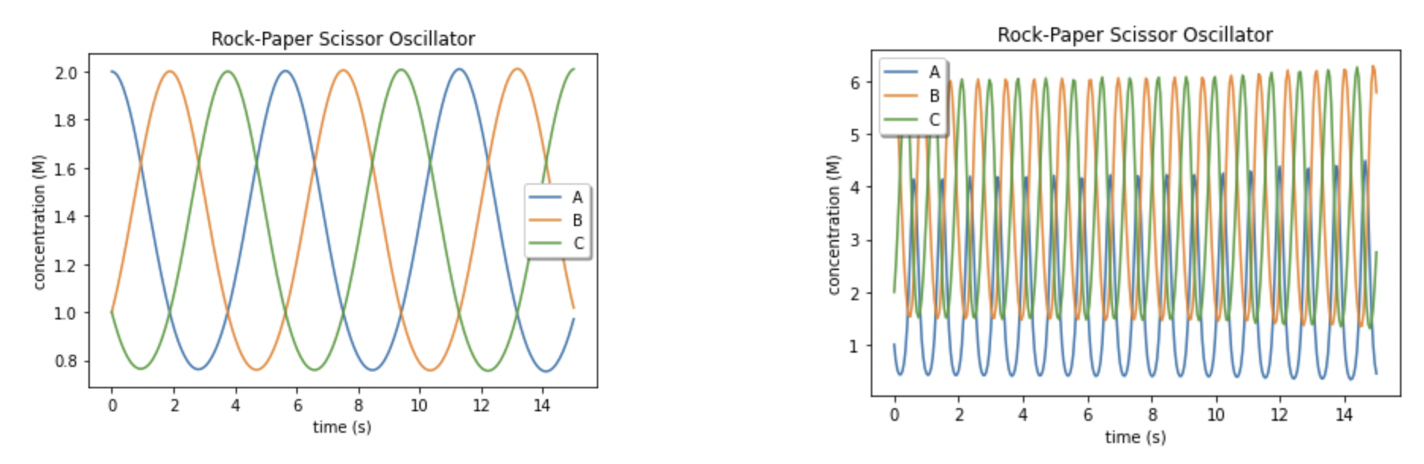
Figure 5. Varying the rates lead to very distinct oscillatory behavior. A. With rates set as the following: k1 = 1, k2 = 1, k3 = 1, we see that the concentrations of species A, B and C are all similar at different times. B. Set the rates to be: k1 = 2, k2 = 1, k3 = 2. Changing rates k1 and k3 dramatically changed the period and the amplitudes of the concentration curves for species A, B, and C.
The mechanics of the RPS Oscillator are described by beginning with a DNA strand that interacts with a fuel DNA molecule. This causes TMSD to be initiated, and the output of this will be a new DNA complex and an incumbent strand which detaches from the original fuel molecule. The new DNA complex will interact with another fuel molecule, and this same process will continue to cyclically occur, one iteration of which is shown in Figure 6.
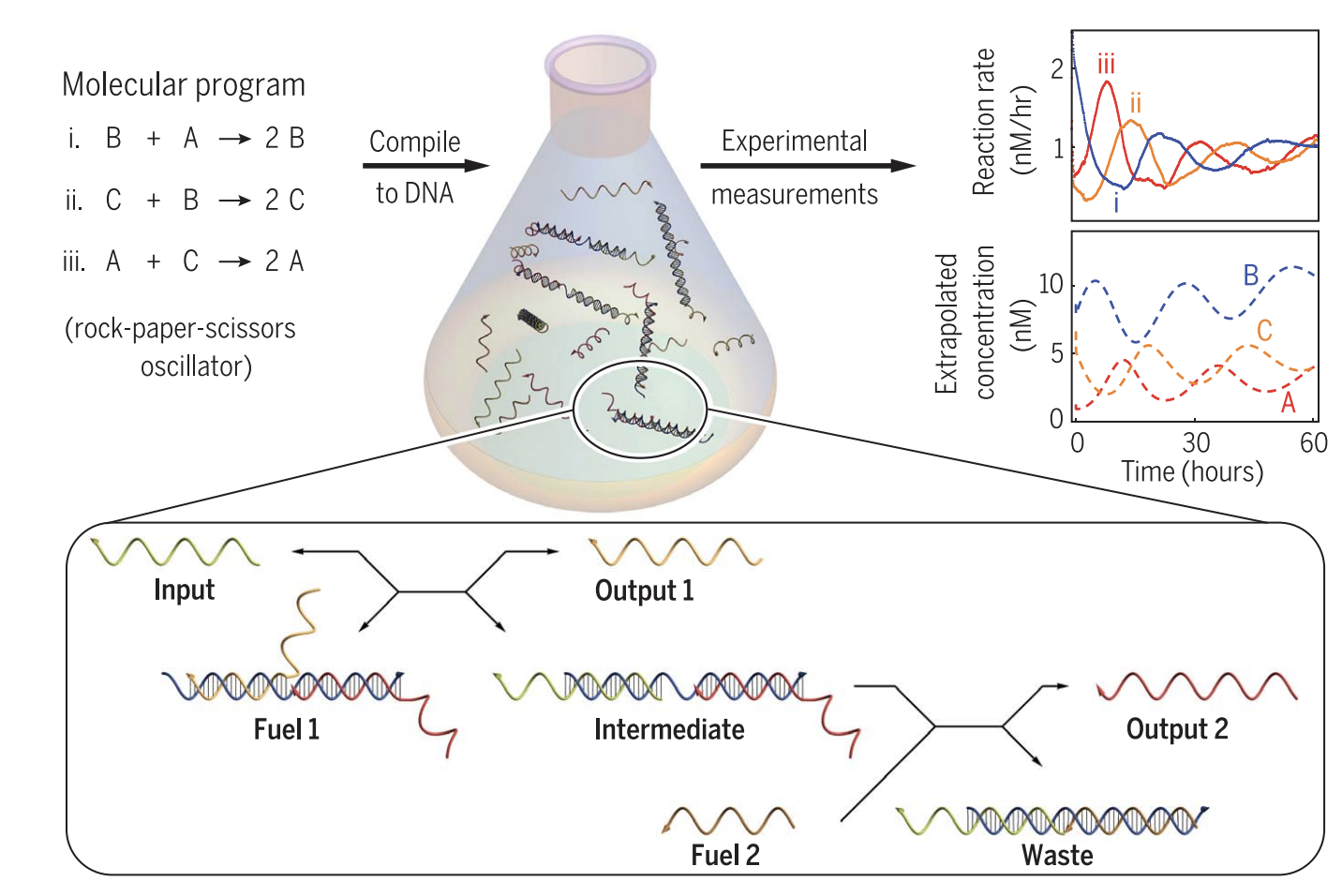
Figure 6. Using DNA to simulate an RPS Oscillator. This diagram only demonstrates what occurs for one reaction module (i.e. X1 + X2 → 2X1). Since two TMSD reactions occur in this module and there are 3 reaction modules per oscillation, for an entire RPS Oscillation, 6 TMSD reactions will occur. Reprinted with permission from “Enzyme-free nucleic acid dynamical systems” by N. Srinivas, 2017, Science.
Since the DNA equivalent of a RPS Oscillation requires 18 total reactions (6 TMSD reactions), and fuel molecules, when the concentration of species A, B and C are plotted at different fuel concentrations (900 nM, 600 nM), we notice that the larger the initial fuel concentration, the faster the oscillations decay (Figure 7).
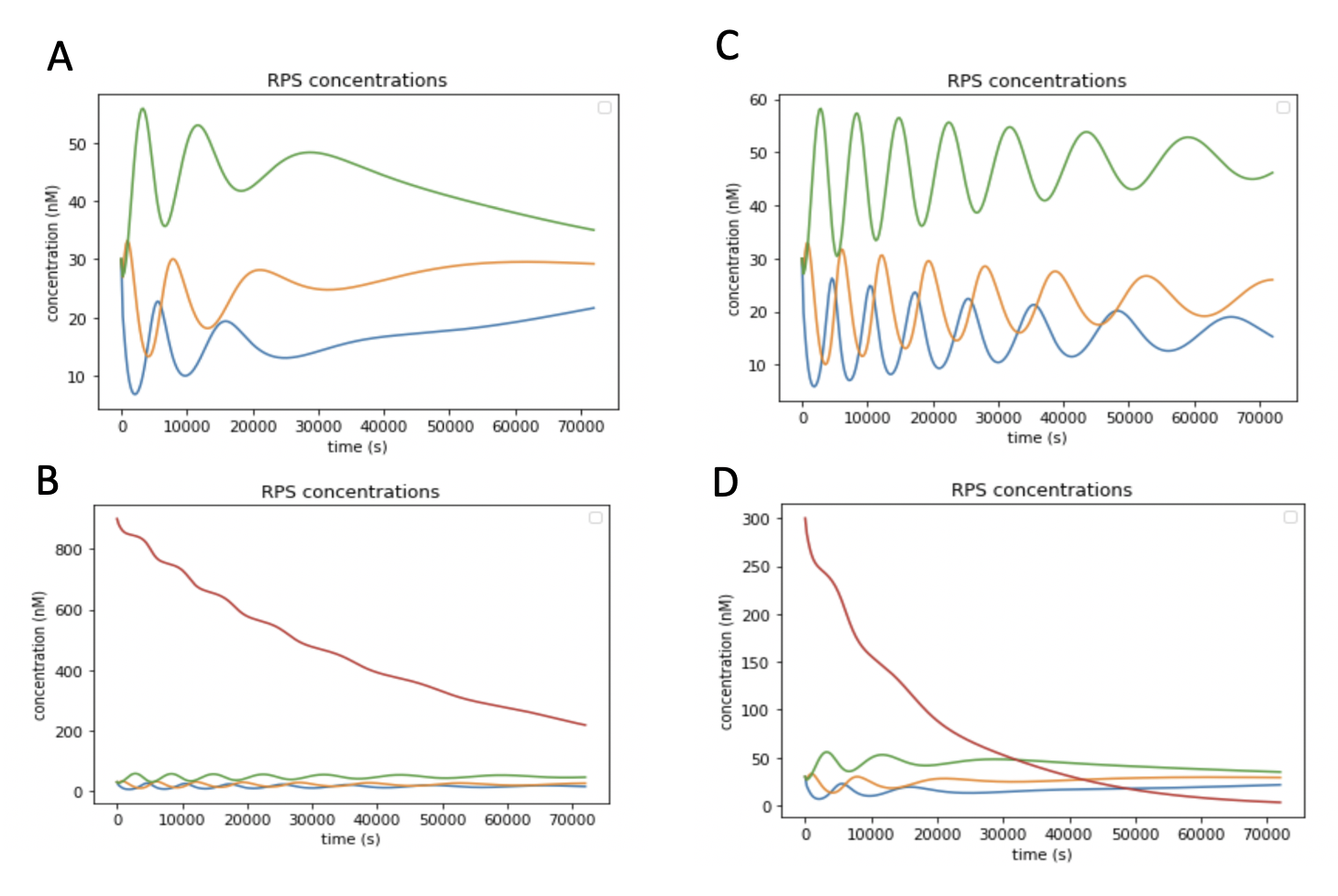
Figure 7. A. The plot of species A, B, and C when the fuel concentration starts at 900 nM. B. Plot from Figure 7A with the fuel concentration over time (red line) added. C. The plots of species A, B, and C when the fuel concentration starts at 300 nM. D. Plot from Figure 7C with the fuel concentration over time (red line) added.
Notice that the plot demonstrates how consumption of fuel molecules (the red line) is what causes species molecules to eventually plateau, rather than noise.
Next, we focus on determining how the rate of an oscillation is dependent on the rate of a TMSD reaction, which is dependent on binding energy and toehold length. First, we identified how the rate of an individual reaction module affects the overall rate. To do so, we varied the values of k1, k2, and k3, and identified their impact on the rates of the corresponding reaction module (Figure 8).

Figure 8. Resulting plots when Ao = 2, Bo = 1, Co = 1, and k1 = 1, k2 = 0.5, k3 = 0.5 unless they are plotted.
A single reaction module is dependent on 6 reaction rates. So, we needed to depict how the overall reaction rate changes based on its sub-rates. We noticed that for larger values of k1, the period of the overall RPS reaction became smaller (Figure 9).
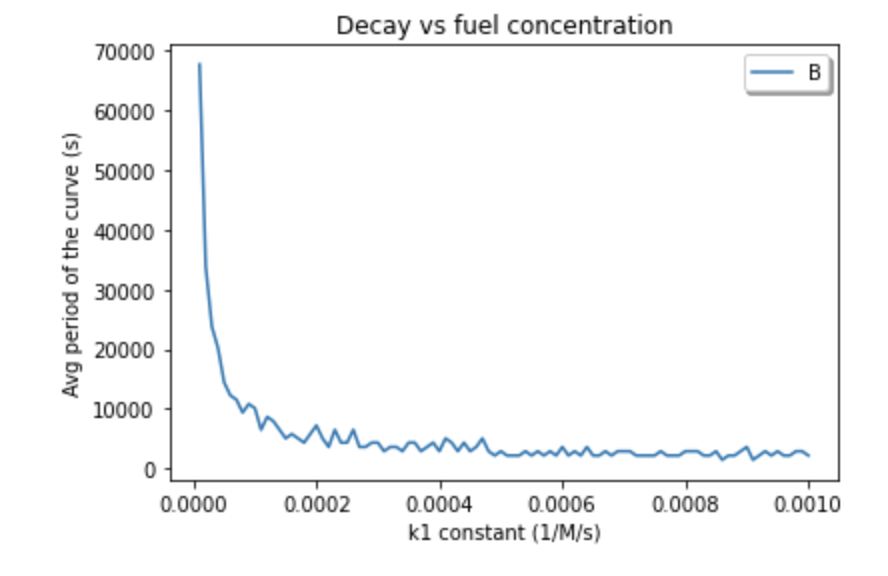
Figure 9. Average period for varying values of k1, the first of the six reaction rates that dictate a single module.
The clear relationship between the period of these oscillations and k1 suggests that the rate of an oscillation must depend on the overall rate of a TMSD reaction and, consequently, the binding energies and strand length.
Discussion
From this project, we reached a few critical outcomes. Most importantly, we were able to successfully model the rate of TMSD as a function of toehold length, while also treating the strand displacement step as a random walk – a procedure which previous literature had failed to account for. We were also able to model how the rates of a rudimentary oscillator could vary by different parameters (i.e. toehold length and binding energies).
Insights from our study of the rate constants governing RPS oscillators will be important for continuing to develop the magnetic handshake system. The methodology we developed to characterize these oscillations can be adapted for various other dynamical phenomena as well. We can construct more complex oscillatory systems, or even use TMSD reactions to simulate various other DNA-like behaviors.
Future work includes building each input in a RPS Oscillator using the magnetic panel sequence. Assuming that the binding sequence and length of our magnetic system are known, we can use the findings outlined to determine the desired rate and create an oscillation. It is important to recognize, though, that there is still room to better identify the overall rate of an oscillation as a function of an individual TMSD reaction. Our methodology did not yield an explicit relationship between the overall oscillation rate and the binding energy. Instead, we characterized the overall oscillatory rate as a function of another oscillatory rate, which is—in turn—dependent on the TMSD rate. Additionally, there are more parameters and rates to account for in the future, since the present goal was to identify the simplest oscillation possible.
Regardless, the outcomes of this project serve as a fundamental approach to assessing more complex DNA behavior, which has many long-term applications. Understanding basic DNA characteristics is crucial for constructing a fully functional magnetic panel system in the future. The development of such a system would not only revamp the way modern-day nanotechnology is both viewed and utilized, but also pave the way for creating new molecular machines and information-storage technology.
Acknowledgements
First and foremost, thank you to Professor Michael Brenner for bringing me on board this summer. The opportunity to work with a lab, attend group meetings and observe research was an invaluable experience. Second, thank you to Dr. Chrisy Xiyu Du, Dr. Agnese Curatolo, and Ofer Kimchi for serving as willing mentors and taking time throughout the week to guide and present me with countless ideas for my project. Third, I would like to thank Professor Erik Winfree for serving as my on-campus mentor and monitoring the progress of my project throughout.
Last but not least, thank you to the California Institute of Technology and Harvard University School of Applied Sciences and Engineering for sponsoring my project this summer and providing the opportunity to conduct research.
References
- Niu, R., Du,C., Esposito, E., Ng, J., Brenner, M., McEuen, P., Cohen, I. (2019). Magnetic handshake materials as a scale-invariant platform for programmed self-assembly. PNAS, 116 (49), 24402-24407. https://www.pnas.org/content/116/49/24402
- Soloveichik, D., Seelig,G., Winfree., E. (2010). DNA as a universal substrate for chemical kinetics. PNAS, 107 (12), 5393-5398; https://www.pnas.org/content/107/12/5393
- Srinivas, N., Ouldridge, T. E., Sulc, P., Schaeffer, J. M., Yurke, B., Louis, A. A., Doye, J. P., & Winfree, E. (2013). On the biophysics and kinetics of toehold-mediated DNA strand displacement. Nucleic acids research, 41(22), 10641–10658. https://doi.org/10.1093/nar/gkt801
- Srinivas, N., Parkin, J., Seelig., G., Winfree., E., Soloveichik., D. (2017). Enzyme-free nucleic acid dynamical systems. Science, 358, ( 6369); https://science.sciencemag.org/content/358/6369/eaal2052
- Zhang, David Yu, and Erik Winfree. “Control of DNA Strand Displacement Kinetics Using Toehold Exchange.” Journal of the American Chemical Society, vol. 131, no. 47, 2009, pp. 17303–17314., doi:10.1021/ja906987s.
